Assays of Therapy by Means of Staphylococcal Bacteriophage
By B. Bruynoghe and J. Maisin.
We have had the opportunity to use the Staphylococcal Bacteriophage for therapeutic purposes, and the few favorable results obtained encourage us to report them. This assay seemed quite logical to us, given that these bacteriolysates contain a principle immediately harmful to Staphylococci. They also contain the staphylococcal antigen essential for any active vaccination, whose therapeutic effects cannot manifest until 5 or 6 days later.
In our previous note, we saw that normal Rabbit serum does not in any way prevent the Bacteriophage from dissolving these microbes. We have also been able to confirm this fact for human serum. For this purpose, we inoculate tubes of sterile serum, on one side with Staphylococci, and on the other side with Bacteriophage and Staphylococci: the inhibition of the latter’s development occurs as it does in broth cultures. The same is true for the dissolution assays of these cultures that have already developed at the moment of the Bacteriophage addition.
Clinical Application and Results
This is not the place to give clinical details concerning the patients we have treated. We are content to report that we applied this therapy to 6 patients suffering from anthrax or furuncles (boils).
We injected doses of bacteriolysates (sterilized by one hour of heating at 56° C or 132.8° F) varying from 0.5 to 2 c.c. as close as possible to the affected region. The effect was not long in manifesting itself by the diminution of the swelling at the lesions and often by the total disappearance of the latter in 24 to 48 hours. Infections that had already reached suppuration (puss formation) empty and dry rapidly.
Following these inoculations, a feverish rise in temperature occurs in some patients; in others, the temperature hardly changes. It seemed to us that this rise in temperature occurs especially in those with vast lesions, where the rapid lysis leads to the resorption of large quantities of microbial products. The injection site is painful and slightly swollen for 24 hours.
Conclusion
We have tried this therapy in patients with anthrax or furuncles, but it’s not impossible that other lesions such as acne and the various staphylococcal complications of other skin conditions could benefit from the same medication.
These observations are obviously not numerous enough to definitively establish the value of this method. They have also not been prolonged enough to determine to what extent these inoculations protect against relapse.
(Institute of Bacteriology of the University of Louvain)
—
Essais de thérapeutique au moyen du Bactériophage du Staphylocoque,
par B. Bruynoghe et J. Maisin.
Nous avons eu l’occasion d’utiliser le Bactériophage du Staphylocoque dans un but thérapeutique et les quelques résultats favorables obtenus nous engagent à les signaler.
Cet essai nous semblait assez logique, étant donné que ces bactériolysats contiennent un principe immédiatement nuisible aux Staphylocoques et l’antigène staphylococcique indispensable à toute vaccination active dont les effets thérapeutiques ne peuvent se manifester que 5 ou 6 jours plus tard.
Dans notre note précédente nous avons vu que le sérum normal du Lapin n’empêche nullement le Bactériophage d’opérer la dissolution de ces microbes. Nous avons pu contrôler également ce fait pour le sérum humain. Dans ce but nous ensemençons des tubes de sérum stérile, d’un côté avec du Staphylocoque, d’un autre côté avec du Bactériophage et du Staphylocoque : l’inhibition sur le développement de ce dernier s’opère comme dans les cultures en bouillon. Il en est de môme pour les essais de dissolution de ces cultures déjà développées au moment de l’addition du Bactériophage.
Ce n’est pas ici la place de donner des détails cliniques concernant les malades que nous avons traités et nous nous contentons de signaler que nous avons appliqué cette thérapeutique chez 6 patients atteints d’anthrax ou de furoncles. Nous avons injecté aussi près que possible de la région malade, des doses de bactériolysats (stérilisées par une heure de chauffage à 56°) variant de 0,5 à 2 c.c. L’effet n’a pas tardé à se manifester par la diminution de l’empâtement au niveau des lésions et souvent par la disparition totale de ces dernières en a/j à 48 heures. Les infections déjà arrivées à la suppuration se vident et sèchent rapidement.
A la suite de ces inoculations, il se produit, chez certains malades, une ascension fébrile, chez d’autres la température ne subit guère de modification. Il nous a semblé que cette élévation de la température se produit surtout chez ceux atteints de vastes lésions et où la lyse rapide entraîne la résorption de grandes quantités de produits microbiens. L’endroit d’injection est durant 24 heures douloureux et légèrement œdématié.
Nous avons essayé cette thérapeutique chez des patients atteints d’anthrax ou de furoncles, mais il n’est pas impossible que d’autres lésions telles que les acmés et les diverses complications staphylococciques d’autres affections cutanées ne puissent profiter de la même médication.
Ces observations ne sont évidemment pas assez nombreuses pour établir définitivement la valeur de cette méthode et elles n’ont pu être assez prolongées pour déterminer jusqu’à quel point ces inoculations protègent contre les rechutes.
(Institut de bactériologie de l’Université de Louvain).
The article was scanned and OCRed using French as the recognition language. The OCRed text was then corrected as needed (as shown in gray). The corrected text was then translated by Claude.ai, which prior to that point was not involved in the effort. Claude.ai was then asked to generate an abstract-style narrative summary (which was then corrected manually as appropriate) and then an assessment of precedence that might be claimed by the work.
Summary by Claude.ai:
This research by R. Appelmans investigated the fate of bacteriophage in animal organisms as part of studies on its therapeutic value. Using guinea pigs and mice, the authors examined bacteriophage behavior following both oral and injectable administration, employing a functional detection method in which organ samples or excreta were added to broth, heated to 56°C to inactivate bacteria while preserving phage, then inoculated with susceptible microbes to detect growth inhibition.
When bacteriophage mixed with bread was given orally to animals, the bacteriophages appeared regularly in feces for several days, persisting longer when susceptible gut microbes were present for it to parasitize. The phage’s resistance to acids and enzymes explained its survival through the digestive tract. However, examination of internal organs revealed no bacteriophage penetration across the intestinal mucosa, a finding the authors attributed to the principle’s inability to dialyze, suggesting it could not pass through biological membranes.
In contrast, injected bacteriophage rapidly entered the bloodstream in accordance with earlier findings by Bordet and Ciuca. However, its presence in blood was brief, as it was progressively eliminated through the kidneys and intestines, disappearing completely within 24 to 48 hours depending on dose, with no traces detected after five days. Notably, the spleen retained substantial quantities of bacteriophage even when it had been cleared from other organs, a phenomenon the authors connected to the spleen’s known role in retaining microbes during infections. This splenic persistence was temporary; after fifteen days, no bacteriophage remained, which the authors attributed to neutralization by antibacteriophage antibodies that form in the organism, as demonstrated by Bordet, Ciuca, and Maisin. The authors emphasized that the bacteriophage detected in organs was not produced by organ-induced bacterial modification as Bordet’s theory might suggest, since control experiments with normal organs or organs from bacteriophage-fed animals never yielded bacteriophage in cultures of susceptible bacteria.
Translation by Claude.ai:
Our research on the therapeutic value of bacteriophage has led us to examine the question of its fate in the organism.
We first investigated whether the bacteriophage supplied to the animal by the digestive route is absorbed. For this purpose, we give guinea pigs and mice the lytic filtrate in question, mixed with bread, and we examine the feces and organs of these animals.
The feces are added to broth, supplemented with a crystal of thymol. After 24 hours we take an ampoule of it, which we heat to 56° for one hour. We place two drops of the contents of this ampoule in a tube of broth, which we inoculate with the receptive microbe.
(Note from claude.ai: “’Réceptif’ (receptive) refers to a bacterial strain susceptible to the particular phage being tested, which is standard terminology for this period.”)
We observe that there is inhibition of growth in these tubes; which we attribute to the presence of bacteriophage and not to the trace of thymol. Indeed, if we heat the contents of this tube to 56° for one hour, to transfer a few drops into a new tube of broth, growth is still absent. According to our research, the bacteriophage principle is regularly found in the feces for several days; it remains there for a longer or shorter time depending on whether or not it finds microbes that it can influence (parasitize). This persistence, moreover, is perfectly explained when one takes into account its resistance to acids and to enzymes, which makes its destruction by digestive juices impossible. (1)
To search for the presence of bacteriophage in the organs, we sacrifice the animals and we aseptically remove the various organs, which we introduce into tubes of broth. If they remain sterile for 24 hours, we inoculate their contents with the receptive microbe, whose growth we monitor. In these media we detect no trace of bacteriophage. The result also remains negative when we add a few drops of the contents of the preceding tubes, heated to 56° for one hour, to broth. The bacteriophage principle therefore does not cross the intestinal mucosa; this fact is perhaps to be related to its inability to dialyze. (2)
As for the bacteriophage injected into animals, here is how we proceeded to determine its fate.
We inject variable doses of it into animals, which we sacrifice after a longer or shorter time, in order to remove the organs and to search in them for the presence of the lytic principle following the technique described above.
From the first hours following these injections, the bacteriophage is absorbed to pass into the blood, in accordance with the data of Bordet and Ciuca (3). However, its stay there is hardly long, for it is progressively eliminated from the organism, by the kidneys and the intestine, to the point of disappearing completely after 24 to 48 hours. The duration of stay varies somewhat with the inoculated dose, but after five days we have never found any trace of it. At this moment however, the spleen still contains notable quantities of it, as can be seen in the table below. This fact is to be related, it seems to us, to the role that this organ plays in infections, where…
(1) Depoorter and Maisin. Archives internationales de Pharmacodynamie et Thérapie, vol. XXV, fasc. V, VI.
(2) C. R. de la Soc. de biol., 26 January 1921.
(3) C. R. de la Soc. de biol., 29 January 1921
…it also intervenes actively to retain the microbes. In the spleen, however, the bacteriophage does not persist, given that fifteen days later, it is completely devoid of it. At this moment, the disappearance results, in our opinion, from the neutralization brought about by the antibacteriophage that forms in the organism, as shown by the experiments of Bordet, Ciuca and Maisin.
Here are the results provided by guinea pigs injected with 0.6 c.c. of d’Herelle’s bacteriophage: [see table image below]
[Table footnote:] Note. – + + Signifies abundant growth as in the control tube: no bacteriophage. – + Signifies considerably less growth than in the control tube: therefore presence of bacteriophage. – Signifies absence of all growth: abundant presence of bacteriophage.
We wish to point out that the bacteriophage that we thus find in the organs is not the product of a modification operated by these organs on the microbe to make it, in accordance with Bordet’s theory, capable of secreting the ferment in question; for this principle was always absent in the tubes of broth, where the microbes receptive to the bacteriophage had undergone contact with normal organs or with organs of animals fed with bacteriophage.
Conclusions: The bacteriophage is not absorbed under normal conditions by the digestive route.
When injected, it passes rapidly into the blood to be eliminated in the urine and feces.
It persists however in the spleen, until the moment when one can admit that the antibacteriophages intervene, to neutralize and destroy it there.
Possible Precedence as suggested by Claude.ai:
[Note: This assessment of precedence is based on the document itself and may not reflect comprehensive historical research.]
This article appears to be an important early systematic study of bacteriophage pharmacokinetics and biodistribution – essentially asking “what happens to phage when you put it in an animal body?” This was a critical question for the therapeutic use of bacteriophage, which was being actively explored in the early 1920s.
Several aspects suggest precedence or importance:
Oral vs. Injectable Routes: The finding that orally administered phage does NOT cross the intestinal barrier was significant for therapeutic applications. This would have important implications for how phage therapy should be administered.
First Detailed Tracking Study: While the authors cite Bordet and Ciuca (1921) for the observation that injected phage enters the blood, Appelmans appears to provide the first detailed tracking of phage through multiple organs over time, including the kinetics of elimination.
Splenic Sequestration: The observation that the spleen retains phage longer than other organs was novel and connected phage behavior to the spleen’s known immunological functions.
Antibody Neutralization: Linking the disappearance of phage from the spleen to antibacteriophage (neutralizing antibodies) helped establish that the immune system recognizes and responds to phage as foreign material.
The dates are telling: the references are from January 1921, suggesting this work was conducted very shortly after (or contemporaneously with) Bordet’s group’s initial observations. Given that Félix d’Herelle only clearly described bacteriophage in 1917, this 1921 work represents research within just a few years of phage discovery, making it part of the foundational literature on phage biology and a key early study informing phage therapy attempts.
OCR Work and Resulting Translation:
Our research on the therapeutic value of bacteriophage has led us to examine the question of its fate in the organism.
We first investigated whether the bacteriophage supplied to the animal by the digestive route is absorbed. For this purpose, we give guinea pigs and mice the lytic filtrate in question, mixed with bread, and we examine the feces and organs of these animals.
The feces are added to broth, supplemented with a crystal of thymol. After 24 hours we take an ampoule of it, which we heat to 56° for one hour. We place two drops of the contents of this ampoule in a tube of broth, which we inoculate with the receptive microbe.
(Note from claude.ai: “’Réceptif’ (receptive) refers to a bacterial strain susceptible to the particular phage being tested, which is standard terminology for this period.”)
LE BACTÉRIOPHAGE DANS L’ORGANISME.
Note de R. Appelmans, présentée par R. Bruynoghe.
Nos recherches sur la valeur thérapeutique du bacteriophage nous ont amené à examiner la question de son sort dans l’organisme.
Nous avons recherché d’abord si le bactériophage fourni à l’animal par voie digestive se résorbe. A cet effet, nous donnons à des Cobayes et des Souris, le filtrate lytique en question, mélangé à du pain et nous examinons les selles et les organes de ces animaux.
Les selles sont ajoutées à du bouillon, additionné d’un cristal de thymol. Après 24 heures nous en prélevons une ampoule, que nous chauffons à 56° pendant une heure. Nous mettons deux goutttes du contenu de cette ampoule dans un tube de bouillon, que nous ensemençons avec le microbe réceptif.
We observe that there is inhibition of growth in these tubes; which we attribute to the presence of bacteriophage and not to the trace of thymol. Indeed, if we heat the contents of this tube to 56° for one hour, to transfer a few drops into a new tube of broth, growth is still absent. According to our research, the bacteriophage principle is regularly found in the feces for several days; it remains there for a longer or shorter time depending on whether or not it finds microbes that it can influence (parasitize). This persistence, moreover, is perfectly explained when one takes into account its resistance to acids and to enzymes, which makes its destruction by digestive juices impossible. (1)
Nous constatons qu’il y a inhibition du développement dans ces tubes ; ce que nous attribuons à la presence de bacteriophage et non à la trace de thymol. En effet, si nous chauffons le contenu de ce tube à 56° pendant une heure, pour en porter quelques gouttes dans un nouveau tube de bouillon, le développement y fait encore défaut. D’après nos recherches le principe bactériophage se retrouve régulièrement dans les selles durant quelques jours ; il y reste plus ou moins longtemps suivant qu’il y trouve ou non des microbes qu’il peut influencer (parasiter). Cette persistance d’ailleurs s’explique parfaitement, quand on tient compte de sa résistance aux acides et aux ferments, qui rend sa destruction par les sucs digestifs impossible. (2)
To search for the presence of bacteriophage in the organs, we sacrifice the animals and we aseptically remove the various organs, which we introduce into tubes of broth. If they remain sterile for 24 hours, we inoculate their contents with the receptive microbe, whose growth we monitor. In these media we detect no trace of bacteriophage. The result also remains negative when we add a few drops of the contents of the preceding tubes, heated to 56° for one hour, to broth. The bacteriophage principle therefore does not cross the intestinal mucosa; this fact is perhaps to be related to its inability to dialyze. (2)
(Note from claude.ai: “The reference to dialysis at the end refers to the inability of bacteriophage particles to pass through dialysis membranes, which was an important observation about their physical properties in early phage research.”)
Pour rechercher la présence du bactériophage dans les organes, nous sacrifions les animaux et nous prélevons aseptiquement les divers organes, que nous introduisons dans les tubes de bouillon. S’ils restent stériles pendant 24 heures, nous ensemençons leur contenu avec le microbe réceptif, dont nous surveillons le développement. Dans ces milieux nous ne décelons pas trace de bactériophage. Le résultat reste également négatif, quand nous ajoutons quelques gouttes du contenu des tubes précédents, chauffés à 56° pendant une heure, à du bouillon. Le principe bactériophage ne franchit donc pas la muqueuse intestinale ; ce fait est peut-être à rapprocher de son inaptitude à la dialyse. (2)
As for the bacteriophage injected into animals, here is how we proceeded to determine its fate.
Quant aux bactériophage injecté aux animaux, voici comment nous avons procédé pour en déterminer le sort.
We inject variable doses of it into animals, which we sacrifice after a longer or shorter time, in order to remove the organs and to search in them for the presence of the lytic principle following the technique described above.
Nous en injectons des doses variables à des animaux, que nous sacrifions après un temps plus ou moins long, afin de prélever les organes et d’y rechercher la présence du principe lytique suivant la technique exposée plus haut.
From the first hours following these injections, the bacteriophage is absorbed to pass into the blood, in accordance with the data of Bordet and Ciuca (3). However, its stay there is hardly long, for it is progressively eliminated from the organism, by the kidneys and the intestine, to the point of disappearing completely after 24 to 48 hours. The duration of stay varies somewhat with the inoculated dose, but after five days we have never found any trace of it. At this moment however, the spleen still contains notable quantities of it, as can be seen in the table below. This fact is to be related, it seems to us, to the role that this organ plays in infections, where…
(1) Depoorter and Maisin. Archives internationales de Pharmacodynamie et Thérapie, vol. XXV, fasc. V, VI.
(2) C. R. de la Soc. de biol., 26 January 1921.
(3) C. R. de la Soc. de biol., 29 January 1921
Dès les premières heures qui suivent ces injections, le bacteriophage se résorbe pour passer dans le sang, conformément aux données de Bordet et Ciuca (3). Toutefois son séjour n’y est guère long, car il s’élimine progressivement de l’organisme, par les reins et l’intestin, au point de disparaître complètement au bout de 24 à 48 heures. La durée du séjour est quelque peu variable avec la dose inoculée, mais après cinq jours nous n’en avons plus jamais trouvé trace. A ce moment toutefois, la rate en contient encore des quantités notables, ainsi qu’on peut le voir dans le tableau ci-dessous. Ce fait est à rapprocher, nous semble-t-il, du rôle que cet organe joue dans les infections, où…
(1) Depoorler el Maisin. Archives internationales de Pharmacodynamie et Thérapie, vol. XXV, fase. V, VI.
(2) C. R. de la Soc. de bioi’.,26 janvier 1921.
(3) C. R. de la Soc. de biol., 29 janvier 1921
…it also intervenes actively to retain the microbes. In the spleen, however, the bacteriophage does not persist, given that fifteen days later, it is completely devoid of it. At this moment, the disappearance results, in our opinion, from the neutralization brought about by the antibacteriophage that forms in the organism, as shown by the experiments of Bordet, Ciuca and Maisin.
…il intervient également activement pour retenir les microbes. Dans la rate le bactériophage ne persiste toutefois pas, étant donné que quinze jours après, elle en est tout à fait dépourvue. A ce moment, la disparition résulte à notre avis, de la neutralisation opérée par l’antibactériophage qui se forme dans l’organisme, ainsi qu’il résulte des expériences de Bordet, Ciuca et Maisin.
Here are the results provided by guinea pigs injected with 0.6 c.c. of d’Herelle’s bacteriophage: [see table image below]
[Table footnote:] Note. – + + Signifies abundant growth as in the control tube: no bacteriophage. – + Signifies considerably less growth than in the control tube: therefore presence of bacteriophage. – Signifies absence of all growth: abundant presence of bacteriophage.
Voici les résultats fournis par des Cobayes injectés avec 0,6 c.c. du bactériophage de d’Herelle :
[These are words from the table:] Foie, Rein, Rale, Coeur, Sang, tué, Urines, Testicule, Poumon, Contrôle d’enemencement.
[This is the table’s footnote:] Nota. – + + Signifie développement abondant comme dans Ie tube Contrôle : pas de bactériophage. – + Signifie déyeloppement considérablement moindre que da’ns le tube contrôle : done présence de bactériophage. – Signifie absence de lout dévelopment : présence abondante de bactériophage.
We wish to point out that the bacteriophage that we thus find in the organs is not the product of a modification operated by these organs on the microbe to make it, in accordance with Bordet’s theory, capable of secreting the ferment in question; for this principle was always absent in the tubes of broth, where the microbes receptive to the bacteriophage had undergone contact with normal organs or with organs of animals fed with bacteriophage.
Conclusions: The bacteriophage is not absorbed under normal conditions by the digestive route.
When injected, it passes rapidly into the blood to be eliminated in the urine and feces.
It persists however in the spleen, until the moment when one can admit that the antibacteriophages intervene, to neutralize and destroy it there.
Nous tenons à faire remarquer que le bactériophage que nous trouvons ainsi dans les organes, n’est pas le produit d’une modification opérée pas ces organes sur le microbe pour le rendre, conformément à la théorie de Bordet, apte à secréter le ferment en question ; car ce principe faisait toujours défaut dans les tubes de bouillon, où les microbes réceptifs. au bacteriophage avaient subi le contact d’organes normaux ou d’organes d’animaux nourris avec du bactériophage.
Conclusions : Le bactériophage ne se résorbe pas dans les conditions normales par voie digestive.
Injecté, il passe rapidement dans le sang pour s’éliminer dans l’urine et les selles.
Il persiste toutefois dans la rate, jusqu’au moment où on peut admettre que les antibactériophages interviennent, pour l’y neutraliser et l’y détruire.
In a Preliminary Note [Comptes rendus, t. 165, 1917, p. 373] I described a filtering microbe found in the excreta of convalescents from bacillary dysentery. The use of a technique less perfect than the one I had used at first [Comptes rendus du la SociétédeBiologie, séance du 7 décembre 1918] combined with the systematic examination of the stools of thirty-four patients, all suffering from dysentery due to Shiga bacilli, and several of whom could be followed daily from the beginning of the illness until the end of convalescence, allowed me to study in a more complete manner the mode of action of the bacteriophage microbe and to specify its role in the evolution of the disease.
In cases of bacillary dysentery, even very serious ones, but in which the patient’s condition improves rapidly, the bacteriophage microbe manifests its presence in a very active manner from the outset, both on cultures of the bacillus isolated from the patient’s excrement and on laboratory Shiga strains, from the moment when the symptoms begin to improve. The bacteriophage power with respect to the dysentery bacillus abruptly ceases to be detectable at the beginning of convalescence. From this moment on, repeated examinations also show the absence of pathogenic bacilli.
In cases where the disease is prolonged, the bacteriophage microbe shows only a nonexistent or slightly marked action, as long as the patient’s condition remains stationary. If, in some cases, the bactericidal action is relatively high on strains which have undergone numerous passages on culture media, on the other hand, it is always inappreciable or very weak on cultures of the bacillus coming from the patient under observation. The improvement is manifested as soon as the bacteriophage action becomes energetic with respect to the latter.
In long-term and relapsing forms, the bacteriophage power of the filtering microbe may, at certain times, be very energetic with respect to cultured bacilli and variable from one day to the next, although always relatively weak, with respect to the patient’s bacillus. Recovery closely follows the moment when the action of the bacteriophage microbe manifests itself in an equally intense manner for both strains. This action persists, with fluctuations in activity, as long as the patient remains a carrier of germs. This latter fact would even be likely to facilitate the detection of germ carriers, the detection of the bacteriophage microbe being simpler and more reliable than the search for the pathogenic bacillus in the stools.
I was able to verify that the action of the bacteriophage microbe was preponderant, not only with regard to the disappearance of the dysentery bacillus from the intestine once the disease had declared itself, but also during its outbreak. During the recent epidemic, I had the opportunity to observe several extremely mild cases in which the symptoms were limited to a few sputum and two or three diarrheal stools: now, in all these cases, the bacteriophage microbe was, from the beginning, present and endowed with a high antagonistic power. Despite the benignity of the affection, it was indeed dysentery because, in three of these cases, I was able to isolate from the first diarrheal stool emitted a typical Shiga bacillus.
The bacteriophage microbe pre-exists in the intestine where it normally lives at the expense of B. coli. In normal stools, its antagonistic power towards the latter bacillus is always very weak; it can become considerable in various morbid states, in certain forms of enteritis and common diarrhea, for example. The presence of dysenteric bacilli in the intestine first determines a considerable exaltation of the virulence of the bacteriophage microbe towards B. coli, then, by a more or less rapid habituation, this virulence is exalted towards the dysenteric bacillus; it immediately or gradually reaches a considerable power leading to the rapid or gradual disappearance of the pathogenic bacillus. If the virulence of the bacteriophage microbe is immediately exalted [“very active”?], the dysenteric bacilli are destroyed from the start of their culture in the intestinal contents, the disease aborts before any symptoms or is limited to a few temporary disorders. If, for a cause which remains to be determined, the virulence of the bacteriophage microbe with respect to the pathogenic microbe does not manifest itself immediately or only manifests itself weakly, a fight is established between the two organisms, the dysentery bacilli multiply in the intestinal contents, infiltrate the mucous membrane, the disease breaks out and the patient’s condition then faithfully records the fluctuations in the fight. In summary, the pathogenesis and pathology of bacillary dysentery are dominated by two opposing factors: the dysentery bacillus, the pathogenic agent, and the filtering microbe bacteriophage, the agent of immunity. As a corollary, the experiment on the rabbit shows that cultures of the bacteriophage microbe have a preventive and curative power in the experimental disease; on the other hand, the bacteriophage microbe is invariably present in the intestine of the sick as soon as the symptoms improve; it therefore seems logical to propose, as a treatment for bacillary dysentery, the administration, as soon as the first symptoms appear, of active cultures of the bacteriophage microbe.
¶ In a note published in the Society of Biology on November 19, 1919, d’Hérelle indicated a method for measuring Bacteriophage. He puts 1/50,000 teaspoon of Bacteriophage into 10 c.c. of an emulsion containing approximately 250 million microbes per teaspoon and spreads a scoop of this mixture (i.e. 1/100 c.c.) onto a slanted agar tube. He then counts the number of dissolution areas and estimates from this the number of ultramicrobes. In fact, the Bacteriophage does not act like a liquid, exerting the same action over the entire seeding surface; lysis only occurs at locations where an ultramicrobe has been deposited on the agar.
¶ This method is, however, rather difficult to carry out because the clarification ranges [plaques?] can be extremely small and therefore not very apparent. Let us add to this that, when they are somewhat large, we could suppose that some of them result from a lack of seeding at this level [I’m unsure what this means].
¶ For the dosage, we tried the process of successive dilutions recommended by Miquel for water analysis. For this purpose we introduce, into tubes seeded with the microbe capable of undergoing lysis, decreasing quantities of Bacteriophage 1/10, 1/100, 1/1000, up to 1/1,000,000,000,000 [that’s 1/1 trillion or 1/1012!], according to that the microbe has normal development or undergoes immediate or subsequent lysis we admit the presence or the absence of the Bacteriophage in question.
¶ When we dose the Bacteriophage coming from tubes, inoculated with the same number of drops of Bacteriophage and the same number of drops of culture, we constant on both sides the same drops of culture [this too is phrasing I struggle with], we note on both sides the same activity. However, it happens from time to time that a given dilution provides a negative seeding for Bacteriophage, while the dilution ten times stronger still contains some. This is explained quite well with the notion of Bacteriophage presuming a living being. We also observe similar facts in the counting of water germs using Miquel’s method.
¶ This technique allowed us to make the following observations:
¶ 1. The Bacteriophage introduced into broth inoculated with lysable microbes, increases quantitatively during the stay in the oven [I assume this is an oven incubator]. This increase is approximately the same for a Bacteriophage and a given culture, whether or not the seeding was massive with the lysable culture or with the Bacteriophage, whether there was immediate inhibition on development or late lysis.
¶ 2. The centrifugation pellet of the Bacteriophage (8,000 revolutions per minute for one hour) does not contain more active elements than the upper liquid.
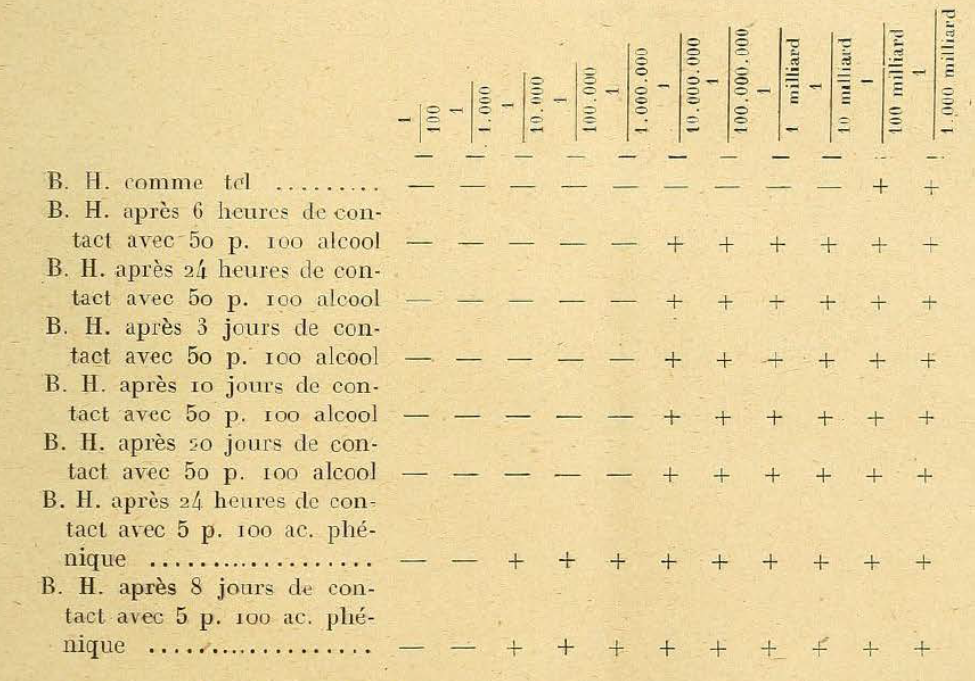
¶ 3. Certain chemical influences, which apparently leave the Bacteriophage intact, nevertheless considerably reduce its activity when the effect produced by these agents is examined by the dosage method. So when we put the Bacteriophage in contact with alcohol at 50 p. 100 or with 5 p. carbolic acid. 100 reviews massively we find [I don’t understand this phrasing]. In these filtrates, Bacteriophage is still active, but with the dosage method we see that the content is considerably reduced, as can be seen from the attached table:
¶ Note. — B. H. means Bacteriophage d’Herelle; — means absence of development of the microbe and presence of Bacteriophage; + means normal development of microbes and absence of Bacteriophage. Fact worthy of being noted: under the influence of alcohol and carbolic acid, the reduction of Bacteriophage occurs quickly, without being progressive: it stops at a certain level, as if the filtrate contained a certain number of elements more resistant to the harmful agents in question.
¶ (Bacteriology Laboratory of the University of Louvain).
An important question for those with ecological and evolutionary biological interests is when a given situation is of ecological relevance vs. when it might be more of evolutionary biological importance (Abedon, 2022b). Telling the difference can be important for all of us.
Ecology by definition is the interaction of organisms with their environments.
We can describe phage use as antibacterial agents, that is, phage therapy, as an example of community ecology, or more precisely an applied community ecology. This is community ecology because there is more than one species of organism involved, i.e., as making up an ecological community. Minimally this is the phage (species #1) and the targeted bacterium (species #2), but also of importance is the treated body (species #3).
By definition, bacterial resistance to phages is ecological, as it describes a specific type of interaction, in this case between at least two species, the phage and the bacterium. That the resistance ‘interaction’ is one of ‘non-‘ or ‘less-‘ contact by the bacterium with the phage antagonist is only a detail, just so long as this lack of interaction is phenotypic, i.e., as opposed to the phage and bacterium instead just happening to exist in different places.
Bacterial resistance to phages also of course can have evolutionary aspects.
Evolution by definition is a change in allele frequencies in at least one species, or, more precisely, changes in allele frequencies in one population, in either case as observed over time.
Often the changes in allele frequency that we care most about are consequences of the impact of natural selection, and natural selection under most circumstances has a strong ecological component. Indeed, natural selection in most cases can be defined as the impact of ecology on evolutionary biology (and hence, as an aside, the existence of the science of evolutionary ecology).
By definition yet again, changes in the frequency of phage-resistance alleles within a bacterial population is an evolutionary process and typically these changes are a consequence of natural selection. The selective agent would be that phage population that is negatively affecting a bacterial population, resulting in increases in the frequency of whatever bacterial alleles are conferring protection from this phage.
Of interest to phage therapy is that this ecology driving evolutionary biology can in turn drive ecology. Specifically, once the frequency of alleles conferring phage resistance are high enough within a targeted bacterial population, then the applied community ecology of phage therapy can be affected, e.g., phage therapy can stop working.
In addition, if the frequency of a phage-resisting allele is found to be 100% within the bacterial population, following phage treatment (i.e., a frequency of 1.0), then if nothing else this is indicative that bacterial survival – an ecological issue – in this instance likely is a function of the occurrence of phage resistance.
If the frequency of phage resistance instead is 0% following phage treatment (0.0), then if nothing else this is indicative that bacterial survival (again, an ecological issue) was not a function of the occurrence of phage resistance. In fact, any frequency of phage resistance below 100% within a targeted bacterial population means that phage-sensitive bacteria are persisting despite phage treatment. For an example of the latter, see, e.g., Box 2 of Abedon (2022c).
Phage-resistant bacteria may display reduced virulence against bodies or may be subsequently treated with a different phage. Consequently, in some ways phage-resistant bacteria are not necessarily that big of a deal as a midpoint of a phage treatment, and this can be particularly if a diversity of other treatment phages are available. Phage resistance is not desired nor welcome, of course, but evolution of phage resistance also is not a certain indication of phage therapy microbiological failure.
That, by the way, to a degree contrasts with the evolution of antibiotic resistance that can occur over the course of antibiotic treatments, which can indeed be associated with treatment failures with high likelihood. One difference is something called antagonistic pleiotropy – not to be confused with antagonistic coevolution (Abedon, 2022a)!!! – i.e., whether or not resistance alleles are otherwise costly to the carrying organism (Abedon, 2022d). If resistance is both easily attained and not ecologically costly, then, well, that can be problematic, particularly given only mono therapies. Another difference is the sheer abundance of diverse, typically safe-to-use phages that often can be available to phage therapists (Abedon and Thomas-Abedon, 2010).
In any case, the persistence of phage-sensitive bacteria despite phage treatment probably means that, for whatever reason, treatment phages are not able to successful infect targeted bacteria despite those bacteria being phage sensitive; again, see Box 2 of Abedon (2022c). This frankly should be viewed as a big deal as essentially by definition it implies a phage therapy microbiological failure, one that may or may not be easily rectified, or at least a lack of complete eradication of phage-sensitive bacteria. I mean, how does one deal with phages not being able to easily reach and/or kill the otherwise phage-sensitive bacteria they are targeting?
Perhaps, as an answer to that question, some other phage that happens to be able to do a better job of reaching and/or killing those otherwise phage-sensitive bacteria might be available for use. That certainly is possible, but at this point in time we really aren’t all that good at figuring out what might constitute a better phage for phage therapy, other than in terms of host range – though see for example Bull et al. (2002; 2019) – and particularly better than the phage that we started with, presumably assuming that the first phage tried we thought was the better phage for phage therapy, hence why it was used first.
Of course, we almost take it as a matter of faith that phage carriage of extracellular polymeric substance (EPS) depolymerases (Danis-Wlodarczyk et al., 2021b) will solve many problems of phage penetration to targeted bacteria. But whether that is actually true in all instances, e.g., such as phage distribution throughout lungs – yet again, see Box 2 of Abedon (2022c) – is in my opinion just not known.
How might use of a phage cocktail instead result in complete eradication of a phage sensitive bacteria when a monophage does not? Just better odds that at least one of the phages used will be particularly good at achieving this? As another aside (Danis-Wlodarczyk et al., 2021a; Abedon, 2022c), note that it can be helpful to just apply a phage or phages at higher or multiple doses before giving up on a given treatment strategy!
At any rate, not being able to eradicate bacteria from an infection even though those bacteria are sensitive to a given treatment can be a far greater problem than failure that get rid of bacteria that explicitly are not susceptible to a treatment protocol. That is, there exits a basic problem in the applied ecology of treatments if not even phage-sensitive bacteria can be removed in full, just as there is a basic problem for an antibiotic treatment if the antibiotic is unable to fully eliminate even antibiotic-sensitive bacteria, a.k.a., the concept of antibiotic tolerance. For a bit on the latter, see Appendix A1 of in fact yet yet again, Abedon (2022c).
So where exactly am I going with this? At the end of a phage treatment, it is important to know whether the frequency of phage-resistant bacteria among the targeted bacterial population is high (at or approaching 100%) rather than low (near 0%). But high precision in that measurement, e.g., more than just whole percentage-point differences, really is not all that important. Why not?
Especially ecologically, there likely is little difference between 0.1%, 0.01%, or even maybe 10% or 50% of the bacteria being phage resistant, as that will still leave an awful lot of phage-sensitive bacteria having escaped phages during treatment. At some probably higher frequency of phage resistance we might come to feel that the frequency of remaining phage-sensitive bacteria is less important, but exactly where that point lies is difficult to say. My gut feeling, though, is that at the point where we start having to do statistics to tell the difference, i.e., at a point where higher precision in measurements becomes important, the importance of differences in the frequencies of phage-resistant bacteria – 100% or a tiny bit less than 100% – probably are no longer all that relevant. (For consideration of the statistics of plating-based enumeration, see Abedon and Katsaounis, 2021.)
Ah, you are saying, clearly therefore I am leading up to claiming that if we are interested instead in the evolutionary biology phage resistance, then in that case we really should care about measuring resistance frequencies with higher precision. And you would be absolutely right! Except also maybe not.
The problem here is that a key word in the definition of evolution that we are using is “Change”, and by definition change cannot be measured using only a single data point, or in the case of quantifying evolutionary change, a single time point. Thus, no matter how precisely you measure the endpoint frequency of phage-resistant bacteria, that will not tell you that evolution has occurred in the course of phage therapy treatment, much less how much evolution.
Here is the basis of this latter point: At the start of an experiment, if your population of bacteria ever is going to contain phage-resistant members, then it likely already does contain those mutants (this, by the way, is why only-qualitative determinations that phage resistance is present, e.g., such as following phage treatments, are basically meaningless). Exceptional would be if the starting number of bacteria involved is so low that this number is, e.g., less than the inverse of the rate of mutation to phage resistance. Thus, for every time a bacterium divides, let’s say that there is a probability of 10-5 that a mutation to phage-resistance will occur. If so, then in a population of 106 bacteria, on average 10 phage-resistant bacteria will be expected to be present, more or less (Abedon et al., 2021).
That last part, “More or less”, is crucial, however, as the frequency with which mutations conferring phage resistance are expected to be present is predicted to somewhat “Fluctuate” about an average (Luria and Delbrück, 1943). In practice, this means that even if you precisely know bacterial rates of mutation to resistance to a given phage, you still will not know how many phage-resistant bacterial mutants will be present prior to the start of treatments. (As yet another aside, actually calculating mutation rates, vs. just mutant frequencies, is a not trivial thing to do.)
Without knowing the frequency of phage resistance prior to the start of treatments, then you are only really guessing whether evolution has occurred in the course of a phage treatment, no matter how precisely frequencies of phage resistance may be measured after a treatment is done.
In short, ecologically, the precision of measures of frequencies of bacterial phage resistance need not be all that high to possess high value in understanding the outcome of phage treatments. I mean, either phage-sensitive bacteria have persisted despite prior treatments or they have not, without a need to describe percentages with precision past the decimal point. Thus, 50.0% vs. 50.1%? Who cares? Indeed, 50% vs. 51%, who cares?
Alternatively, if one really cares about being precise in monitoring the evolution of phage resistance, then the most important place to emphasize that precision actually should be prior to the start of treatments, i.e., prior to initial phage application, and only then should one be measuring frequencies of phage resistance after treatments as well. But don’t forget that you need to have this information for explicitly that bacterial culture that is being treated, since evolutionarily all we really will care about is how a specific bacterial culture as a population changes in allele frequencies over time, and in phage therapy that bacterial population is precisely the one that you are treating.
Even so, how much more than order-of-magnitude precision do we really need in monitoring the evolution of phage resistance during phage treatments? Will we really care for example if the frequency of phage-resistant bacteria have changed from 10-5 to 10-5.5? And how hard would we have to try to be sure that such a relatively small change is actually real? I mean, seriously, except for the most hard-core evolutionary experiments, who would really care?
For what it is worth, when I look at the outcome of a phage treatment, if all of the targeted bacteria remaining are phage resistant, then I know what went wrong (clue: the bacteria have evolved resistance to the treatment phages, i.e., an evolutionary outcome). But when I look at the outcome of a phage therapy experiment and a substantial portion of the bacteria remaining are still phage sensitive, then more often than not I can only speculate as to what might have gone wrong, except again for those bacteria that have evolved phage resistance (Abedon, 2022c). Still, this latter scenario should be viewed at least as an ecologically relevant outcome.
But bottom line: Obtaining an additional decimal place or two in describing the frequency of phage-resisting alleles within the treated bacterial population generally will not greatly aid in improving the precision of our applied ecological speculation.
Afterthought:
You would think that this essay came into existence as a natural outgrowth of the cited publications, particularly Abedon (2022c), but you would be wrong! On the other hand, I did wait a few months until that publication was published and available open access. Thanks for your interest!
Literature Cited:
Abedon, S. T. 2022a. A primer on phage-bacterium antagonistic coevolution, p. 293-315. In Bacteriophages as Drivers of Evolution: An Evolutionary Ecological Perspective. Springer, Cham, Switzerland. https://link.springer.com/chapter/10.1007/978-3-030-94309-7_25
Abedon, S. T. 2022b. Frequency-dependent selection in light of phage exposure, p. 275-292. In Bacteriophages as Drivers of Evolution: An Evolutionary Ecological Perspective. Springer, Cham, Switzerland. https://link.springer.com/chapter/10.1007/978-3-030-94309-7_24
Abedon, S. T., and T. I. Katsaounis. 2021. Detection of bacteriophages: statistical aspects of plaque assay, p. 539-560. In D. Harper, S. T. Abedon, B. H. Burrowes, and M. McConville (ed.), Bacteriophages: Biology, Technology, Therapy. Springer Nature Switzerland AG, New York City. https://link.springer.com/referenceworkentry/10.1007/978-3-319-40598-8_17-1
Abedon, S. T., K. M. Danis-Wlodarczyk, and D. J. Wozniak. 2021. Phage cocktail development for bacteriophage therapy: toward improving spectrum of activity breadth and depth. Pharmaceuticals 14:1019. https://pubmed.ncbi.nlm.nih.gov/34681243/
Bull, J. J., B. R. Levin, T. DeRouin, N. Walker, and C. A. Bloch. 2002. Dynamics of success and failure in phage and antibiotic therapy in experimental infections. BMC Microbiol. 2:35. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC138797/
Bull, J. J., B. R. Levin, and I. J. Molineux. 2019. Promises and pitfalls of in vivo evolution to improve phage therapy. Viruses 11:1083. https://pubmed.ncbi.nlm.nih.gov/31766537/
Danis-Wlodarczyk, K., K. Dabrowska, and S. T. Abedon. 2021a. Phage therapy: the pharmacology of antibacterial viruses. Curr. Issues Mol. Biol. 40:81-164. https://pubmed.ncbi.nlm.nih.gov/32503951/
Danis-Wlodarczyk, K. M., D. J. Wozniak, and S. T. Abedon. 2021b. Treating bacterial infections with bacteriophage-based enzybiotics: in vitro, in vivo and clinical application. Antibiotics 10:1497. https://pubmed.ncbi.nlm.nih.gov/34943709/
Luria, S. E., and M. Delbrück. 1943. Mutations of bacteria from virus sensitivity to virus resistance. Genetics 28:491-511. https://pubmed.ncbi.nlm.nih.gov/17247100/
Here is a something worth knowing about, from Pires, D.P., H. Oliveira, L.D. Melo, S. Sillankorva, and J. Azeredo. 2016. Bacteriophage-encoded depolymerases: their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 100:2141-2151. [PubMed], (calls to figure and table excluded from quote):
Based on our search, the huge majority of phage depolymerases (126 proteins) are encoded in the same open reading frame of phage structural proteins (mostly on tail fibers, base plates, but sometimes also in the neck) or in close proximity to those genes, and were thus considered as structural proteins. Twenty other depolymerases found in this work might be soluble proteins since they are distant from any structural gene.
Depolymerases that are only soluble, that is, not virion attached, presumably are only useful in the immediate vicinity of phage-lysed bacteria, e.g., towards phage burrowing more deeply into biofilms. This perhaps means that phages don’t need depolymerases to initially infect biofilm bacteria (see here for that argument). Depolymerases that are associated with virions, by contrast, presumably are useful as well upon initial phage encounter with a biofilm bacterium.
That the majority of depolymerases are may not be soluble, but instead appear to be associated with virions, is suggestive that depolymerases are employed for the sake of initial encounter between virions and biofilm bacteteria. But this then begs the question of why more phages don’t encoded depolymerases?
Is it that we have trouble recognizing them in sequence data? Is it that bacteria are just too diverse in terms of extracellular polymers produced? (In addition to limiting utility, the latter may also interfere with our ability to detect depolymerase phenotypes during phage growth as plaques.) Is it because for the most part phages can infect biofilm bacteria sufficiently even without depolymerases? Or are there unexplored trade-offs associated with depolymerase encoding, perhaps especially when they are present as structural components of phage virions?
In the three previous paragraphs I am drawing on a tiny bit of past thought as to the role of depolymerases in phage interaction with biofilms, as can be found in my 2011 book, Bacteriophages and Biofilms. In particular, from p. 23 (of the revised pagination version, or p. 27 of the original… don’t ask…):
Ecologically, EPS depolymerases improve phage movement that occurs either adjacent to or distant from a phage’s parental infection. If distant, then movement towards bacteria will be enhanced by physical linkage between virions and depolymerases. Alternatively, for more localized movement, then soluble depolymerases may suffice, such as for phage dissemination out of biofilms [2004]. Scholl et al. [2005] thus found that efficiency of plating (EOP) was low for phages encoding a soluble EPS depolymerase when infecting a K1 capsule-producing strain and that an isogenic phage not encoding the depolymerase is “unable to form plaques on lawns of this strain” (p. 4872). This result is suggestive that though initiation of plaques occurred with low efficiency, once those infections commenced then subsequent EPS depolymerization presumably facilitated phage migration towards adjacent bacteria to complete plaque formation. In circumstances where enzymes may not be directly supplied, it should thus be advantageous for those enzymes to be carried by virion particles, if only to increase the efficiency of initial infection. That is, it should be advantageous to phages for enzymes to be present at the point of phage adsorption, by being virion attached, rather than present only immediately following the lyses of phage-infected bacteria [I then illustrate this argument with a figure…].
This quote is from Lee Watkins and J. W. Costerton (1984). Growth and biocide resistance of bacterial biofilms in industrial systems. Chemical Times and Trends (October):35-40.
The article has nothing to do with viruses. The quote is interesting, however, since it speaks to the question of how exactly to employ viruses in the biocontrol of microorganisms, specifically in the biocontrol and indeed elimination of biofilms from surfaces (quotation marks in the original):
lt is important to be able to answer that old question “Shall we slug with a biocide, shall we continuously treat with a biocide or shall we soak with a biocide—what is the best deal for the situation?”
To me these three alternatives are distinguishable in terms of how we might think about treatment of bacteria or biofilms with phages, that is, phage-mediated biocontrol of bacteria, or phage therapy.
The first alternative I interpret as the application of large amounts of biocide over short periods, perhaps in a single dose, i.e., slugging, or what we might describe as passive treatment in the case of phage therapy. Keep in mind that passive treatment should mean that for every bacterium targeted not only should at least 10 phages be added but at least 10 phages should be adsorbing, per adsorbable bacterium.
Continuous application, to me, would imply the application of lower but still minimally effective concentrations of biocide over longer periods. Continuous application represents an extreme of multiple dosing, i.e., where the time gap between applications is reduced to zero. Key here is that something other than overwhelming amounts of biocide is being applied, what many (unfortunately) would describe as something other than high multiplicities of infection (MOI) in the case of phage application. One can view such continuous application a preventive, or prophylactic.
Lastly there is soaking, which could also be viewed as continuous application, though this is an application that takes place over a relatively short period, i.e., days or weeks rather than months or years. This would be equivalent to the application of phages by soaking bandages, soaking various absorbent material (one sees mention of “tampons” in various places in the phage therapy literature, though it’s important to realize that the word has a medical definition), or instead via the application of, e.g., Phage Bioderm (for example, as discussed here).
What’s missing, of course, are any assumptions that the biocide will replicate in situ, i.e., so-called active treatment, which is typically considered to be a hallmark of phage-mediated biocontrol/phage therapy. That absence, though, is not unexpected given that this is from a discussion of chemical and physical anti-biofilm biocides rather than of phages.
Still, it once again is nice to see that there really is little that is new under the sun. When dealing with bacterial infections, particularly chronic ones which are associated with biofilms, it can be important to keep in mind these ideas:
We can hit them very hard (literally overkill) over short periods,
We can hit them less hard (minimally adequate biocide concentrations) but over long periods, perhaps particularly towards prevention, or
We can soak the infections over intermediate periods, presumably with periodic re-invigoration of dosing, using antibacterial levels which, also presumably, are somewhat in excess of what might be viewed as minimally adequate.
Any other approach, unless backed by hard data, should be considered to represent mostly wishful thinking.
Some additional reading:
Abedon, S.T. 2016. Bacteriophage exploitation of bacterial biofilms: phage preference for less mature targets? FEMS Microbiol. Lett. 363:fnv246. [PubMed]
Abedon, S.T. 2016. Commentary: phage therapy of staphylococcal chronic osteomyelitis in experimental animal model. Front. Microbiol. 7:1251. [PubMed]
Abedon, S.T. 2016. Phage therapy dosing: The problem(s) with multiplicity of infection (MOI). Bacteriophage 6:e1220348. [PubMed]
Abedon, S.T. 2014. Bacteriophages as drugs: the pharmacology of phage therapy., p. 69-100. In J. Borysowski, R. Miedzybrodzki, and A. Górski (eds.), Phage Therapy: Current Research and Applications. Caister Academic Press, Norfolk, UK.
Way back in 2010 we (Abedon and Thomas-Abedon) suggested that the growth of phage plaques within bacterial lawns could serve as mimics of bacteriophage interaction with bacterial biofiolms. In fact, we made a rather extensive argument with six Roman numeraled points: (i) constraint of bacterial movement, (ii) bacterial growth within lawns as microcolonies, (iii) inhibition of phage movement, (iv) plaque-like phage growth within actual biofilms, (v) possible temporary shielding of bacteria within lawn microcolonies from phage attack, and (vi) variation in bacterial physiologies again as found within microcolonies within lawns and as potentially equivalent to bacterial microcolonies within biofilms. We concluded that, “Given these similarities, phage plaques as a facile laboratory model therefore could enrich our understanding of phage-bacterial interrelations as they may occur during the phage therapy of biofilm-producing bacterial infections.”
Indeed, we noted as well that Gallet et al. (2009) described phage formation of plaques also as phage growth within a “biofilm-like environment”.
Here I provide a quote from an earlier publication which serves to further these arguments. From Gilbert and Brown (1995) [Mechanisms of the protection of bacterial biofilms from antibacterial agents, p. 118-130. In J. W. Costerton and H. Lappin-Scott (ed.), Microbial biofilms. Cambridge University Press, Cambridge, UK.], p. 119:
The most simple in vitro method of generating biofilms to study antimicrobial sensitivity is to inoculate the surface of an agar plate to produce a confluent growth. Such cultures, whilst not fully duplicating the in vivo situation, have been suggested to model the close proximity of individual cells to one another and the various gradients found in biofilms. In this respect, colonies grown on agar may bе representative of biofilms at solid-air interfaces.
Of course, one cannot claim that bacteria growing within soft agar overlays are perfect representations of naturally occurring biofilm structures. Nonetheless, as we’ve noted previously, e.g., Abedon and Yin (2009), plaque formation within them is a lot more complex than people otherwise may realize.
Phage tails, to the extent that they display smaller diameters than phage virions as a whole, might contribute to nonenzymatic virion translocation into EPS [Extracellular Polymeric Substance, i.e., as associated with biofilm matrix], perhaps with longer, narrower tails permitting deeper or faster local penetration to biofilm-surface located bacteria.
Perhaps not unexpectedly, I now find that this was not an entirely original thought. From Wilkinson (1958), p. 68:
Physical blocking of the surface receptor. Can the presence of a capsule protect the cell simply because of its physical properties? Presumably an infective phage must be able to inject its DNA through the cytoplasmic membrane and, therefore, the main body of the phage must be at a distance from the cytoplasmic membrane smaller than the length of the phage tail (rarely longer than 150 mµ [meaning 150 nm]). Therefore, any layer outside the cytoplasmic membrane which is greater in thickness than 150 mµ and is impermeable to phages will act as a nonspecific phage inhibitor. It has already been shown that a capsule is by definition greater than 150 mµ in thickness and that it is probably impermeable to particles of the size of a phage head (about 100 mµ). An additional barrier to the phage might be the high negative charge of the polysaccharide capsular surface. In confirmation of this role, capsulate bacteria have been reported to be generally phage resistant…
In my defense, my suggestion pointed specifically to longer phage tails, and the general thrust of my article was that it is especially less mature aspects of biofilms which may be more vulnerable to phages. Less maturity, in other words, might be associated with less thick biofilm matrix, e.g., as perhaps associated with new growth on biofilm surfaces.
Indeed, Wilkinson discusses further an article, unfortunately which is not in English, suggesting that when polymeric substance material is thinner then successful phage adsorption may be more likely (as continuing directly from the previous quote):
Thus, Kauffmann and Vahlne (82) found that most capsulate strains of E. coli were resistant to phage and that when capsulate strains were attacked, there was a proportionality between the thickness of the capsule and the resistance.
One could speculate therefore that it could be, conversely, that shorter tails would be less able to penetrate thicker “capsulate”.
It should be noted that I am making no claims that phage tails exist solely for the sake of penetrating extracellular polymers towards adsorption of bacteria, though it is entirely possible that such penetration could serve as a benefit of possessing especially “longer, narrower tails”.
Myth: Freezing phage T7 can select for deletion mutations…
From Clark and Geary (1973), with emphasis added:
ATCC freeze-dries phages for distribution (Clark and Geary, 1969). Davis and Hyman (1971) have reported the existence of at least two authentically different T7 phage strains, T7M (Meselsohn) and T7L (Luria) (ATCC 11303-B7). They also stated that T7L lyophilized stock from ATCC contained a high percentage of deletions in the DNA molecule, and they attributed this to lyophilization selecting for DNA deletions. However, since this publication, ATCC prevailed upon these authors to test the ATCC broth stock of T7L, which had been maintained at 4° C unlyophilized or unfrozen since its deposit in the Collection by Luria in 1952. They reported in a letter to W. A. Clark that the T7L stock before freeze-drying contained the same deletions as the freeze-dried product.
In other words, the evidence is not consistent with freezing selecting for deletion mutations in phage T7…
REFERENCES
Clark, W. A., and D. Geary. 1969. The collection of bacteriophages at the American Type Culture Collection, p. 179-187. In T. Nei (ed.), Freezing and Drying Microorganisms. University Park Press, Baltimore.
Clark, W. A., and D. Geary. 1973. Preservation of bacteriophages by freezing and freeze-drying. Cryobiology 10:351-360.
Davis, R. W., and R. W. Hyman. 1971. A study of evolution: The DNA base sequence homology between coliphages T3 and T7. J. Mol. Biol. 62:287-301.